Project 13.1

- PhD student: Tom Sistermans
- Supervisor: Susanne Foitzik
- Co-Supervisors: Peter Baumann
- Research Group
My PhD project is focussed around the interaction between the parasitic tapeworm Anomotaenia brevis and the ant Temnothorax nylanderi. As with many parasites, the host’s phenotype is radically different from the uninfected phenotype. We think this is caused by the parasite actively manipulating the host’s gene expression. My task is to find that out.
Parasites represent over half of species on earth (Windsor, 1998). This diversity has made it difficult for biologists to find characteristics, aside from the fact that parasites gain fitness at the cost of their host, that unify parasites. There is one characteristic that seems to unite many parasites, which is their ability to manipulate their host in favour of their own fitness (Thomas et al. 2005; Poulin, 2010). This includes small behavioural changes like human enterobiasis secreting acid, causing an itching sensation in their host where they lay their eggs (Hugot et al. 1999), to the almost complete control of the fungus Ophiocordyceps unilateralis on the nervous system of their insect host (Evans & Samson, 1984).
Still, within this diversity of host manipulation comes an important question about parasitology in general: How is the parasite able to manipulate the host in both simple and complex interactions? Although the answer to this question is going to be unique for every parasite-host interaction, from an evolutionary perspective, one should be able to hypothesise that some of them share common pathways of achieving this. For instance, instead of a parasite continuously secreting proteins to influence host behaviour, a parasite could interfere with the host phenotype on a level of gene expression itself (Saeij et al. 2007; Villares et al. 2020). This could come in especially useful for the parasite when the host has rarely expressed genetic material in place that could assist in parasite fitness.
Likely among the most striking examples of this phenotypic plasticity can be found in social insects, where the phenotype of clones can radically differ depending on the caste of the individual. It is perhaps not surprising that arguably the most successful group of social insects, ants, have such a diversity in host-parasite interactions that can cause radical phenotypic changes (De Bekker et al. 2018). For example, the previously named fungus Ophiocordyceps unilateralis mostly infects ant species (Evans & Samson, 1984). Furthermore, there is the famous example of the ant Cephalotes atratus and the parasitic nematode Myrmeconema neotropicum, in which the nematode causes the ant’s cuticle to display a red colour reminiscent of berries to increase the chance of berry-eating birds to spot them (Poinar & Yanoviak, 2008; De Bekker et al. 2018).
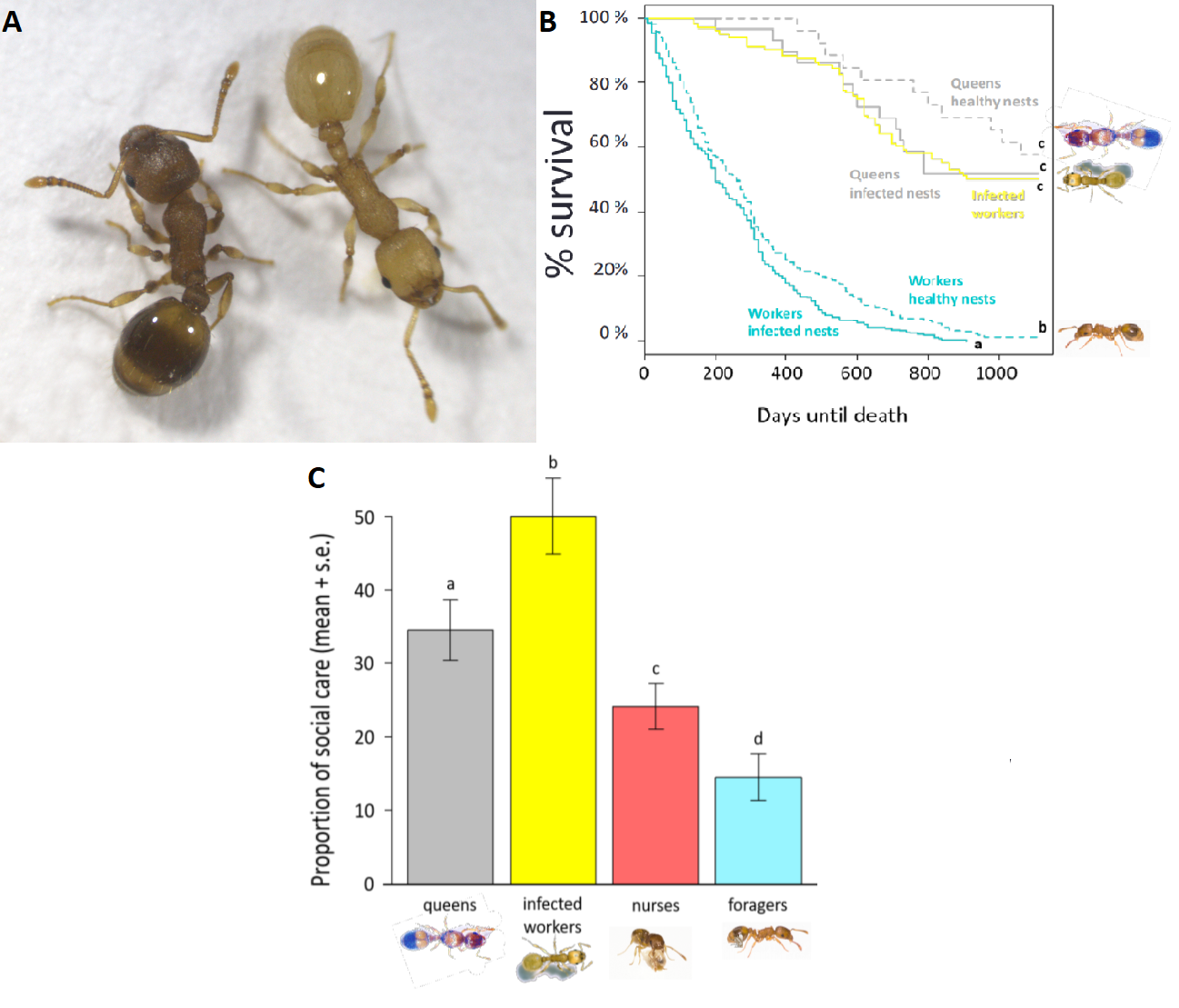
This brings us to the system studied in this PhD, the interaction between the parasitic cestode Anomotaenia brevis and its host Temnothorax nylanderi. Infected T. nylanderi workers have a yellow-coloured cuticle (figure 1a), a smaller head, they remain in the nest as opposed to other workers (Beros et al. 2015) and they have a longer lifespan (Beros et al. unpublished). Nevertheless, these characteristics are not necessarily unique to infected workers. As seen in figure 1b, the long lifespan is very similar to that of T. nylanderi queens. The fact that they remain in the nest and are kept alive by other workers (figure 1c) might be because they display a similar hydrocarbon profile to that of larvae or queens. There is therefore a good reason to suspect that A. brevis does not necessarily invoke changes in phenotype directly, but instead activates or deactivates host genes to its own benefit.